Cellular Biology
course-show.h1-title
Discover the secrets of cell organelle formation and renewal. Studying the biogenesis and dynamics of tissue organelles will help you understand how these structures, essential to cell life, are generated, modified, and maintained over time.
Introduction
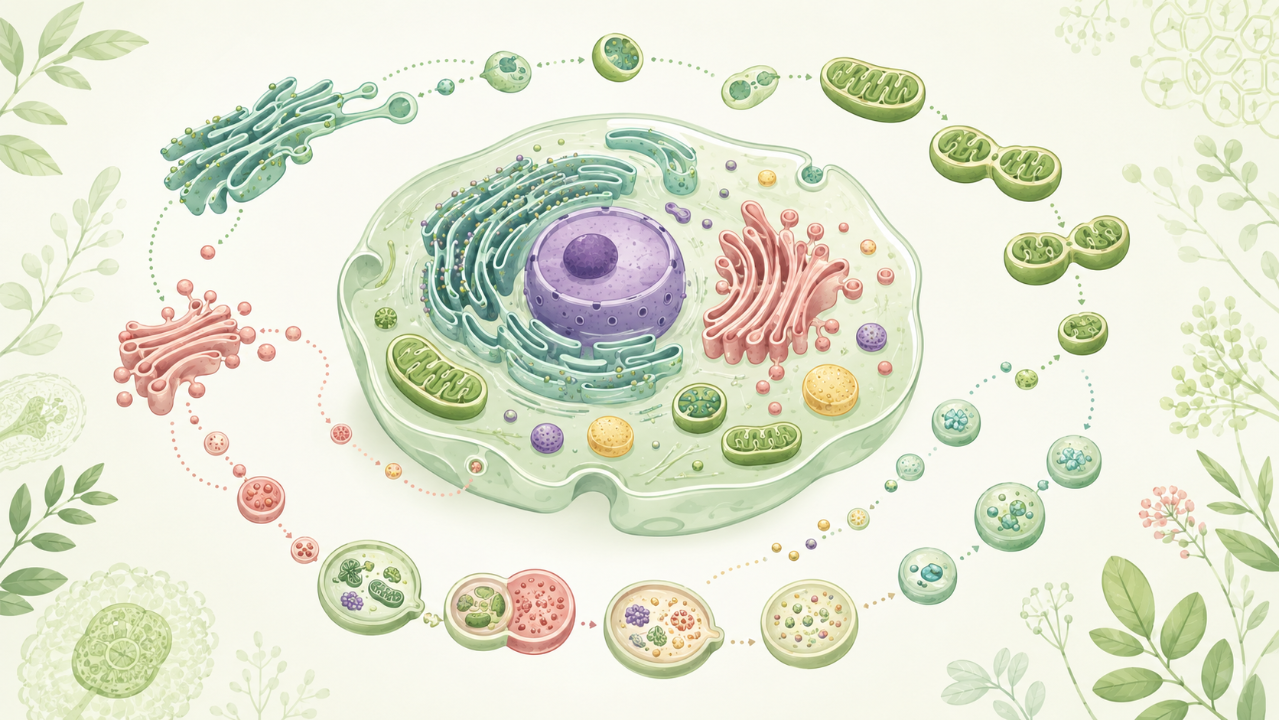
The biogenesis and renewal of organelles is a fundamental aspect of cellular biology that plays a crucial role in maintaining the structural and functional integrity of eukaryotic cells. This process involves the synthesis, assembly, and maintenance of various organelles, including mitochondria, endoplasmic reticulum (ER), Golgi apparatus, lysosomes, peroxisomes, and vacuoles. Understanding these mechanisms is essential for comprehending cellular homeostasis, cell division, and response to stress.
Mitochondria Biogenesis and Renewal
De novo synthesis of mitochondria
Mitochondria are semiautonomous organelles that replicate independently of the nuclear genome. The process of de novo synthesis begins with the transcription and translation of mitochondrial genes, resulting in the production of mitochondrial proteins in the mitochondrial matrix. Subsequently, these proteins form complexes essential for oxidative phosphorylation, which is responsible for ATP production.
Mitochondrial division and fission
The division of mitochondria occurs through a series of events that involve the coordination between the cytoskeleton and the outer mitochondrial membrane. Dynamin-related proteins (DRP1, FIS1, and MID49/MFP1) orchestrate the fission process by constricting the mitochondrion at specific sites, ultimately leading to its division into two daughter organelles.
Mitochondrial renewal
Mitochondrial renewal mainly occurs through a process known as autophagy, where damaged or dysfunctional mitochondria are targeted and engulfed by lysosomes for degradation. The resulting metabolites can then be reutilized to synthesize new mitochondria or provide energy for the cell.
Endoplasmic Reticulum (ER) Biogenesis and Renewal
De novo synthesis of ER
The de novo synthesis of ER involves the coordinated action of several proteins, including SEC61 translocon complexes that facilitate the insertion of proteins into the ER lumen. Additionally, lipids such as phosphatidylcholine and phosphatidylinositol are essential for ER membrane biogenesis.
ER expansion and renewal
ER expansion is mediated by the regulation of calcium levels within the cell, which controls the activity of ER-resident enzymes involved in lipid synthesis. During periods of increased protein folding demand, such as during stress or growth, the ER can undergo expansion by increasing its membrane surface area through budding and branching.
In terms of renewal, ER renewal primarily occurs via a process known as ERAD (Endoplasmic Reticulum-Associated Degradation), where misfolded proteins are targeted for degradation by the proteasome or lysosomes, allowing for the removal and replacement of damaged portions of the ER.
Golgi Apparatus Biogenesis and Renewal
De novo synthesis of Golgi apparatus
The de novo synthesis of the Golgi apparatus begins with the assembly of cisternae from COPI-coated vesicles that bud off from ER exit sites. These vesicles transport proteins destined for the Golgi and are essential for establishing the distinct compartments within the organelle.
Golgi apparatus renewal
Golgi apparatus renewal primarily occurs through a process known as cisternal maturation, where immature cisternae move from the cis-Golgi to the trans-Golgi, acquiring enzymes and modifying proteins along the way. Additionally, damaged or dysfunctional Golgi cisternae can be targeted for degradation by autophagy, allowing for the replacement of these organelles with new ones.
Lysosomes Biogenesis and Renewal
De novo synthesis of lysosomes
The de novo synthesis of lysosomes involves the coalescence of autophagosomes, which engulf cytoplasmic components for degradation. Additionally, lysosome biogenesis requires the targeting and transport of hydrolytic enzymes to the lysosomal membrane via specific transporters such as LIMP-2 (Lysozyme-like protein 2).
Lysosomes renewal
The renewal of lysosomes primarily occurs through autophagy, where damaged or dysfunctional lysosomes are targeted for degradation and replaced with new ones. Additionally, the cell can regulate the number of lysosomes by altering their fusion and fission rates.
Peroxisomes Biogenesis and Renewal
De novo synthesis of peroxisomes
The de novo synthesis of peroxisomes is a complex process that requires the coordinated action of several proteins, including Pex proteins that facilitate the assembly and import of proteins into the organelle. Additionally, the biogenesis of peroxisomes is facilitated by lipids such as phosphatidylcholine and cardiolipin.
Peroxisomes renewal
Peroxisome renewal primarily occurs through a process known as peroxisome division, where the organelle elongates and constricts at specific sites, eventually dividing into two daughter peroxisomes. Additionally, damaged or dysfunctional peroxisomes can be targeted for degradation by autophagy, allowing for their replacement with new ones.
Vacuoles Biogenesis and Renewal
De novo synthesis of vacuoles
The de novo synthesis of vacuoles is primarily observed in plant cells, where it involves the sequestration of water and solutes within a membrane-bound compartment. The formation of vacuoles is regulated by various genes that encode for proteins involved in vesicle trafficking, membrane biogenesis, and osmoregulation.
Vacuoles renewal
Vacuole renewal primarily occurs through the turnover of their contents, where solutes are transported out of the vacuole for reuse or excretion. Additionally, damaged or dysfunctional vacuoles can be targeted for degradation by autophagy, allowing for their replacement with new ones.